Plastoglobules vs Lipid Droplets: Distinct Roles in Senescing Leaves based on JXB Volume 75, Issue 20 – Editor’s choice
Plastoglobules vs Lipid Droplets: Distinct Roles in Senescing Leaves
Discover the JXB Volume 75, Issue 20 – Editor’s choice by Malini Muthu Karpagam. This article highlights the following publication: "Roles of plastoglobules and lipid droplets in leaf neutral lipid accumulation during senescence and nitrogen deprivation”.
This article highlights the following publication:
Roles of plastoglobules and lipid droplets in leaf neutral lipid accumulation during senescence and nitrogen deprivation
Denis Coulon , Houda Nacir , Delphine Bahammou , Juliette Jouhet , Jean-Jacques Bessoule , Laëtitia Fouillen , Claire Bréhélin
Journal of Experimental Botany, Volume 75, Issue 20, 2024, Pages 6542-6562, https://doi.org/10.1093/jxb/erae301
“To die, to be really dead, that must be glorious!” wrote Bela Lugosi, reflecting a distinctly human outlook on the finality of death.
But in plants dying leaves tell a different story. For leaves death is rarely an end, rather, it is more of a metamorphosis. Leaf senescence beautifully epitomizes this paradox through a tightly coordinated dismantling and recycling process during which essential nutrients are salvaged towards actively growing tissues.
Leaf senescence starts in the chloroplast, the photosynthetic machinery of plants. Besides chlorophyll pigments, chloroplasts are also rich in structural lipids like monogalactosyldiacylglycerol (MGDG) and digalactosyldiacylglycerol (DGDG), which constitute the membranes. During natural or stress-induced senescence (such as nitrogen starvation), these structural lipids break down and release free C16:3 and C18:3 fatty acids (FAs). Although these FAs are wealthy energy reservoirs, in excess, they are cell toxic. This necessitates their quick conversion into neutral lipid forms - triacylglycerols (TAGs), steryl-esters (SEs) or phytyl-esters (PEs) to safely store or repurpose them.
These neutral lipids are stored in two distinct compartments: plastoglobules (PGs) and cytosolic lipid droplets (LDs). PGs are small lipid storage bodies in chloroplasts, while LDs are spherical structures that bud from the endoplasmic reticulum and remain in the cytosol. During senescence, MGDG degradation and buildup of neutral lipids is elevated and both compartments increase in abundance. Yet, the precise roles of PGs and LDs and how they coordinate lipid storage logistics remained largely unclear.
In a recent study, Coulon et al. (2024) explored the lipid composition of PGs and LDs in Arabidopsis thaliana leaves under three conditions: natural senescence, nitrogen starvation-induced senescence, and stress recovery (nitrogen resupply). They found that some of the FAs released from MGDG degradation during senescence or nitrogen starvation were incorporated into TAGs, PEs and SEs.
But which neutral lipids are stored in PGs Vs LDs during senescence?
Coulon et al. (2024) found that the types of neutral lipids differ between PGs and LDs. PGs accumulated only a limited amount of TAGs , contradicting the earlier assumption that PGs are major storage sites for TAGs. Instead, PGs stored PEs and surprisingly SEs, that are generally associated only with cytosolic LDs. The cytosolic LDs on the other hand primarily accumulated TAGs during senescence. An even more striking observation was the selective sorting of FAs into PGs and LDs during senescence or nitrogen starvation. To understand this sorting, the authors used Pulse-chase radiolabeling (a method where FA molecules are radiolabeled and tracked over time) along with fad2 mutant analysis. They found that during nitrogen starvation, the C16:3 FAs from MGDG degradation remained in plastids and were esterified and stored in PGs. Meanwhile, the MGDG-released C18:3 FAs were exported from plastids and accumulated as TAGs and SEs in cytosolic LDs. This suggests a selective segregation mechanism, where certain FAs (C16:3) are deliberately retained in the chloroplast while others are sent out for storage or metabolic repurposing.
How do these FAs cross between plastids and LDs?
Using 3D electron tomography, Coulon et al. (2024) showed that the plastid membranes are pressed against LD surfaces, likely forming a "lipid transport bridge" between these two compartments.
The radiolabel tracking also showed that during stress recovery (nitrogen resupply) TAGs were remobilized, possibly towards membrane construction. Meanwhile PEs and SEs were not remobilized, suggesting a TAG-specific remobilization pathway. By exploring this dichotomy in lipid logistics between PGs and LDs, Coulon et al. (2024) open up exciting avenues for understanding the coordinated mechanisms in plants during senescence and stress recovery.
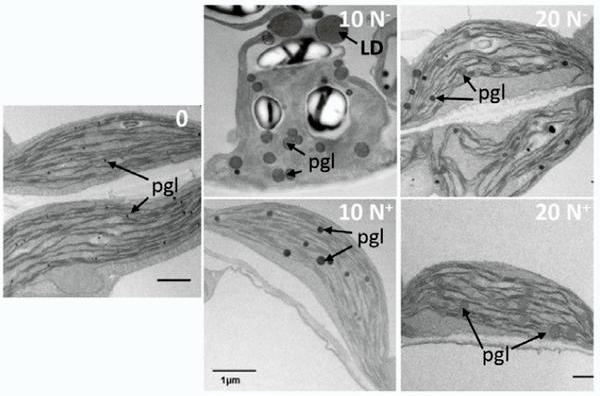
Figure 6B: TEM image shows changes in the ultrastructure in Arabidopsis chloroplasts: under nitrogen starvation (10N–), plastoglobules and lipid droplets increase in number and size, indicating accumulation of neutral lipids. Even after nitrogen is added (20 N+), the plastoglobules do not completely disappear, suggesting incomplete reversal (Figure from Coulon et al., 2024, JXB).
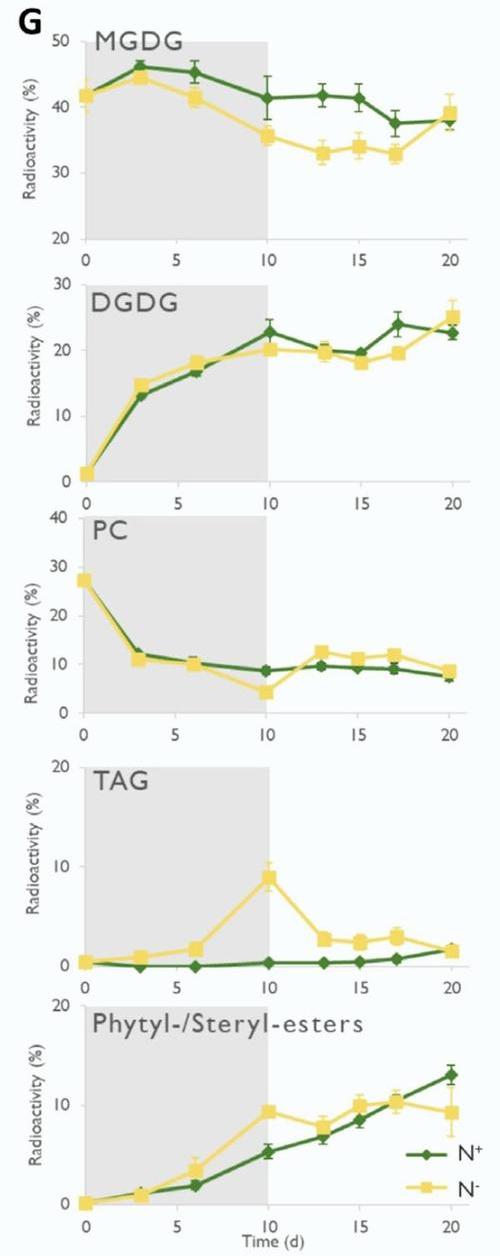
During the recovery phase (white area) in N- seedlings (yellow line), TAGs were remobilized (label decreases), likely rerouted towards membrane building, while PEs/SEs remain unchanged, indicating selective lipid recycling.