October 2025 TPJ Editor choice: Salicylic acid turns down the heat on thermomorphogenesis
Highlighted publications:
Salicylic acid reduces ELF3 phase separation and suppresses thermomorphogenic growth in Arabidopsis
Salicylic acid turns down the heat on thermomorphogenesis
Plants continuously monitor their environment and adjust their growth and physiology to maximise fitness and reproductive success. When exposed to moderately elevated temperatures, they undergo profound morphological changes known as thermomorphogenesis. In the model plant Arabidopsis thaliana, this response is characterised by elongation of hypocotyls and petioles and upward bending of leaves, creating an open architecture that enhances cooling to avoid damaging intracellular temperatures.
The transcription factor PHYTOCHROME INTERACTING FACTOR 4 (PIF4) is central to this process, coordinating multiple phytohormone pathways: PIF4 directly induces expression of auxin biosynthesis genes such as YUCCA 8 (YUC8), generating a mobile auxin signal that moves from the cotyledons and leaves to hypocotyls and petioles. There, auxin triggers local accumulation of brassinosteroids (BRs) and together, auxin and BR signalling cascades stimulate cell elongation. In contrast, the stress-responsive hormones jasmonic acid and abscisic acid dampen thermomorphogenic growth.
Two new studies [MB1] by the groups of Nam-Hai Chua and Danhua Jiang addressed the role of another stress response hormone, salicylic acid (SA), in thermomorphogenesis. Initial observations that osmotic stress represses thermomorphogenesis and upregulates SA biosynthesis, while warm temperature reduces SA content, prompted both groups to investigate the role of SA in thermomorphogenesis more thoroughly. They showed that SA treatment effectively reduced hypocotyl elongation at elevated temperatures (Fig. 1a), an effect that could be mimicked by induced overexpression of the SA biosynthesis gene ISOCHORISMATE SYNTHASE 1 (ICS1) (Fig. 1b). Loss-of-function mutations in ICS1 had the opposite effect, displaying slightly exaggerated elongation at elevated temperatures. Notably, no significant differences were observed at control temperatures, indicating that SA does not act as a general inhibitor of elongation but specifically suppresses high temperature-induced growth.
The authors next asked whether SA affected thermomorphogenesis through PIF4 given its central role in temperature signalling. Elevated temperatures increase PIF4 transcript levels as well as PIF4 protein stability. Mannitol and SA treatments, as well as induced ICS1 overexpression, counteracted the high temperature-induced expression of PIF4 and its downstream target YUC8. A PIF4 overexpression line, in which PIF4 was expressed from the constitutive 35S promoter, was largely insensitive to SA treatment. These findings suggest that SA attenuates thermomorphogenesis mainly by repressing PIF4 at the transcriptional level.
PIF4 transcription is controlled by the temperature sensor EARLY FLOWERING 3 (ELF3), a component of the evening complex. Mutations in elf3 increase hypocotyl elongation, and Song, Chen and colleagues found that this elongation was SA-insensitive. Expression of PIF4 target genes such as YUC8 also stayed high in elf3 after SA or mannitol treatment. Moreover, loss-of-function mutants of ELF4 and LUX ARRYTHMO (LUX), which encode the remaining components of the evening complex, were similarly unresponsive to SA as was elf3. The authors thus concluded that ELF3 constitutes a critical component of SA-dependent repression of thermomorphogenesis.
How does SA affect ELF3’s function? SA treatment and ICS1 overexpression increased ELF3 binding to the PIF4 promoter, while this binding was reduced in an ics1 mutant. ELF3’s activity is controlled by liquid-liquid phase separation, a process by which a uniform solution separates into a dense biomolecular condensate and a dilute phase, mediated by the protein’s prion-like domain. Elevated temperatures promote condensate formation (visible as nuclear speckles of ELF3-mGFP), thereby removing ELF3 from its target genes and inactivating it. SA, mannitol and ICS1 overexpression reduced ELF3 speckle formation in warm conditions (Fig. 1c), whereas speckle formation was enhanced in an ics1 mutant. Interestingly, Chen et al. also demonstrated an effect of SA on ELF3 phase separation in vitro: a purified ELF3 prion-like domain spontaneously formed condensates in vitro (visible as distinct droplets), and the addition of SA suppressed this process.
Chen et al. and Song et al. propose a model in which SA suppresses thermomorphogenesis by reducing ELF3 phase separation, keeping it in its active, DNA-bound form. SA thereby strengthens repression of PIF4 and limits auxin-driven elongation under elevated temperatures. This mechanism appears to be particularly relevant when warm temperatures coincide with additional stresses such as drought, when excessive elongation would increase water loss and be detrimental to plant fitness. SA thus helps plants strike a balance between growth and stress resistance.
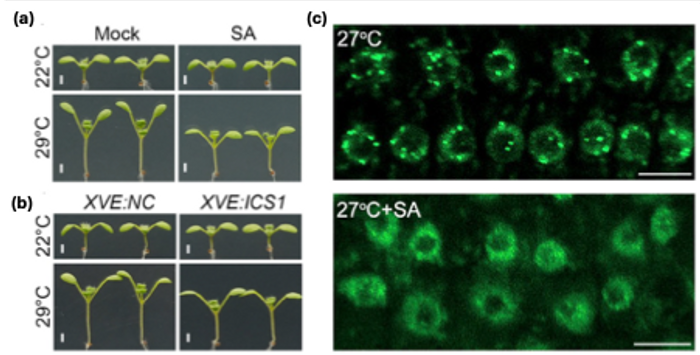
Figure 1. Salicylic acid suppresses thermomorphogenesis via reduced ELF3 phase separation
(a, b) Treatment with SA (a) and β-estradiol-induced expression of ICS1 using the XVE system (b) attenuate hypocotyl elongation at elevated temperatures.
(c) SA treatment reduces the formation of ELF3-mGFP biomolecular condensates (visible as distinct nuclear speckles) at elevated temperatures.
Figure modified from Chen et al. (2025) and Song et al. (2025).




