September 2025 TPJ Editor choice: Riboflavin in phloem sap helps wild tomato combat whiteflies
Riboflavin in phloem sap helps wild tomato combat whiteflies
Highlighting https://doi.org/10.1111/tpj.70363
The whitefly Bemisia tabaci feeds on vegetable and ornamental crops including chili, okra, potato, tobacco and tomato, causing global annual losses up to billions of US dollars. As phloem-feeding insects, B. tabaci feed on host plants by inserting their mouthpart, the stylet, directly into the phloem. During that process, they inject RNA and protein that suppress the plant immune system. Additionally, they can transfer viruses such as the tomato yellow leaf curl virus. Whitefly is a rapidly adapting pest that has evolved resistance to even the most potent chemical treatments. Sustainable whitefly management requires integrating multiple defense layers—including genetic resistance—into comprehensive Integrated Pest Management strategies. Some wild relatives of cultivated tomato are resistant to whitefly damage. Identifying the mechanisms and genes underlying whitefly resistance in wild tomato can equip the vegetable breeding industry with genetic tools to introgress this trait into cultivated varieties.
Most of these resistance mechanisms in wild tomato are based on the production of specialized metabolites in glandular trichomes that are bioactive against B. tabaci. Denkers et al. analysed a defence mechanism that was independent of trichome repellents. The wild tomato accession Solanum chmielewskii was found to be susceptible to adult whiteflies, however, the whiteflies deposited fewer eggs on its leaves and the development of juvenile whitefly stages was hampered. The team hypothesised that this resistance in S. chmielewskii might be due to something in the phloem that specifically targets young stages of whiteflies, the nymphs, which rely on constant feeding from the phloem.
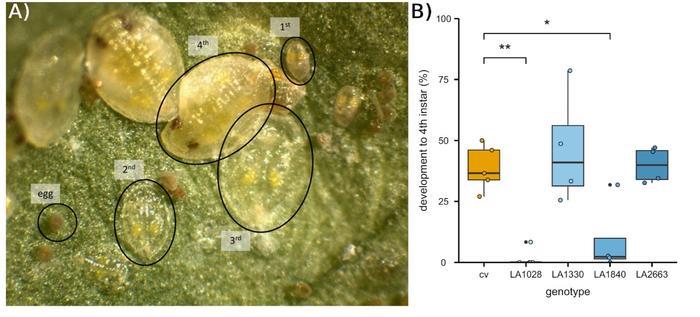
Whiteflies deposit their eggs on host plant leaves, then the first instar nymph hatches and searches for a feeding site towards the leaf vasculature. There, it probes the tissue to access phloem sap, before progressing through three additional nymphal instar stages (Figure 1A). The authors quantified the nymph developmental stages on four S. chmielewskii accessions and on a susceptible tomato cultivar after infestation. Two accessions, LA1028 and LA1840, had fewer eggs deposited and a lower number of nymphs (Figure 1B). To assess whether phloem sap was the causal factor, the authors used LA1840 in grafting experiments. When LA1840 served as a rootstock for the susceptible tomato cultivar shoot, both egg deposition and nymph abundance were reduced, suggesting that the resistance was mediated by a vasculature-mobile factor originating from LA1840.
To identify the metabolite responsible for the resistance, the authors used inbred lines from crosses of LA1840 with a susceptible parent and selected one susceptible and one resistant inbred line for further analysis. To analyze non-volatile compounds in the leaf material, they used Ultra High-Performance Liquid Chromatography-quadrupole Time of Flight and identified 792 features. Then, they applied a Random Forest machine learning approach to identify metabolites associated with resistance. This analysis identified 41 metabolic features as significant discriminators between susceptible and resistant lines. The top candidate was 21-fold more abundant in LA1840 than in the susceptible parental line and was identified as riboflavin.
When riboflavin was supplied to susceptible plants via a hydroponic growth solution, whitefly nymphal development on the leaves was impaired, reproducing the resistance phenotype observed in LA1840. Conversely, treatment of LA1840 plants with a riboflavin synthase inhibitor increased susceptibility, confirming riboflavin as the phloem sap metabolite responsible for resistance against B. tabaci.
The authors propose two possibilities for why riboflavin is a deterrent. First, it might directly inhibit nymphal development when ingested above a threshold concentration, as was demonstrated for aphids. Alternatively, it might act indirectly by triggering other plant defense mechanisms, such as the induction of pathogenesis-related gene expression, as reported in Arabidopsis and tobacco.
|
Figure 1: The wild tomato Solanum chmielewskii is resistant to whitefly infestations. A) Whitefly developmental stages, from egg to first mobile stage to the second, third and fourth immobile stages. B) Compared to the susceptible tomato cultivar (cv), fewer eggs develop to the fourth instar stage on the wild accessions LA1028 and LA1840. Modified from (Denkers et al., 2025). |



